2.4 - Tessuti a
medio contenuto di acqua : muscolo
2.4.1 - Generalita'
Molto concisamente diremo che il tessuto muscolare è suddivisibile
in tre subsistemi:
- muscolatura striata;
- muscolatura liscia;
- muscolatura mista (striata-liscia, relativa al cuore e ad altri organi
interni).
Il primo tipo di muscolatura è quella denominata di tipo "volontaria",
come, ad esempio, quella relativa alla locomozione. La seconda è
quella denominata "involontaria". La maggiore peculiarità delle
cellule muscolari rispetto alle altre cellule è quella di produrre
forza non quando tirata, ma quando posta in compressione. Questo movimento
produce un lavoro meccanico, a spese dell'energia metabolica.
 Fig.24 -La struttura
fibrillare del muscolo
Fig.24 -La struttura
fibrillare del muscolo
In figura 24 si mostra la struttura allungata e composta da diverse
fibre di un muscolo striato. E' evidente la struttura anisotropa del tessuto.
La capacità di contrazione del tessuto è correlata all'attività
metabolica, e comunque alla presenza nel citoplasma di proteine a struttura
filamentosa, contrattili: l'actina e la miosina. A dire il vero queste
proteine, responsabili della creazione della struttura cellulare, sono
presenti in tutte le cellule, ma sono particolarmente abbondanti nel muscolo
ed assumono una struttura a strati. Il lavoro di contrazione avviene grazie
allo scivolamento di strati di actina su strati di miosina in strutture
organizzate dette
miofibrille, ed avviene a spese di scissione di molecole di ATP21
in presenza di ioni calcio.
2.4.2 - Comportamento
dielettrico
Il muscolo è un tessuto particolarmente interessante dal punto
di vista didattico e di ricerca, caratterizzato da un comportamento estremamente
anisotropo, sia per quanto riguarda la costante dielettriche che per la
conducibilità.
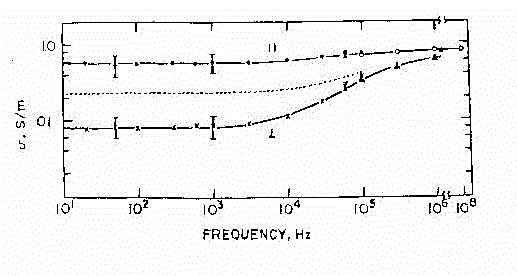 Fig.25
Nella figura 25 abbiamo i risultati di misure sperimentali per la conducibilità
per le due diverse orientazioni del campo ( la linea tratteggiata indica
l'andamento medio, indipendente dall'orientamento del campo ).Si nota la
dipendenza dei dati dalla direzione del campo applicato. Se è parallelo
all'asse più lungo della struttura (indicato in figura con il simbolo
//), avrò una maggiore facilità di spostamento per le cariche
presenti sulla superficie e quindi una conducibilità più
elevata. Si vede poi che aumentando la frequenza del campo applicato, non
si riscontra una sensibile variazione dei valori di s.
Ciò significa che le cariche seguono più agevolmente i cambiamenti
di campo, che i fenomeni di rilassamento si hanno a frequenze maggiori
e creano minore dispersione. Se invece il campo applicato è perpendicolare
all'asse più lungo della struttura (con il simbolo ^
in figura), il movimento per le cariche risulta più complicato e
queste tenderanno a depositarsi sulla superficie della struttura stessa.
In termini quantitativi risulterà per la conducibilità un
valore inizialmente più basso a causa del minor numero di cariche
che riescono a seguire il campo. Il fenomeno di rilassamento che però
si ha, salendo in frequenza, aumenta la conducibilità del sistema,
fino a portarla quasi al livello di quella per il campo applicato parallelamente.
Fig.25
Nella figura 25 abbiamo i risultati di misure sperimentali per la conducibilità
per le due diverse orientazioni del campo ( la linea tratteggiata indica
l'andamento medio, indipendente dall'orientamento del campo ).Si nota la
dipendenza dei dati dalla direzione del campo applicato. Se è parallelo
all'asse più lungo della struttura (indicato in figura con il simbolo
//), avrò una maggiore facilità di spostamento per le cariche
presenti sulla superficie e quindi una conducibilità più
elevata. Si vede poi che aumentando la frequenza del campo applicato, non
si riscontra una sensibile variazione dei valori di s.
Ciò significa che le cariche seguono più agevolmente i cambiamenti
di campo, che i fenomeni di rilassamento si hanno a frequenze maggiori
e creano minore dispersione. Se invece il campo applicato è perpendicolare
all'asse più lungo della struttura (con il simbolo ^
in figura), il movimento per le cariche risulta più complicato e
queste tenderanno a depositarsi sulla superficie della struttura stessa.
In termini quantitativi risulterà per la conducibilità un
valore inizialmente più basso a causa del minor numero di cariche
che riescono a seguire il campo. Il fenomeno di rilassamento che però
si ha, salendo in frequenza, aumenta la conducibilità del sistema,
fino a portarla quasi al livello di quella per il campo applicato parallelamente.
Vediamo ora, per il muscolo, le proprietà dielettriche nelle
tre regioni di maggiore dispersione.
2.4.3 - Dispersione alfa
Una prima teoria supponeva che questo rilassamento traesse origine dalla
polarizzazione dei controioni vicino alla membrana di superficie. Falk
e Fatt fecero però notare che, per giustificare teoricamente i valori
ottenuti sperimentalimente, questo processo avrebbe richiesto densità
di carica fissa sulla superficie troppo elevata per strutture organiche.
Idearono allora un modello che utilizzava i sistemi sarcotubolari rappresentati
in figura 26.
 Fig.26 - Ingrandimento
delle fibre muscolari
Fig.26 - Ingrandimento
delle fibre muscolari
Ipotizzarono che la dispersione fosse dovuta alla bassa impedenza d'ingresso
del sistema. La corrente dunque, alle basse frequenze, scorre all'interno
dei sarcotuboli e la capacità e della resistenza del fluido offerta
dal tessuto risulta aumentata dalla maggiore superficie a disposizione.
Considerando poi un campo E.M. perpendicolare, alle basse frequenze la
permettività era proporzionale alla capacità della membrana,
e la dispersione risultante dal processo di carica del sistema sarcotubolare,
era associata ad un aumento della permettività del tessuto. Risultava,
per il sistema sarcotubolare, una capacità di almeno un ordine di
grandezza maggiore rispetto a quella della membrana esterna. Sperimentalmente
però non sempre c'è stato buon accordo con i dati attesi.
Anche da ciò deriva l'enorme difficoltà a comprovare una
teoria del genere e la supposizione che invece possa esserci l'interferenza
di altri fenomeni.
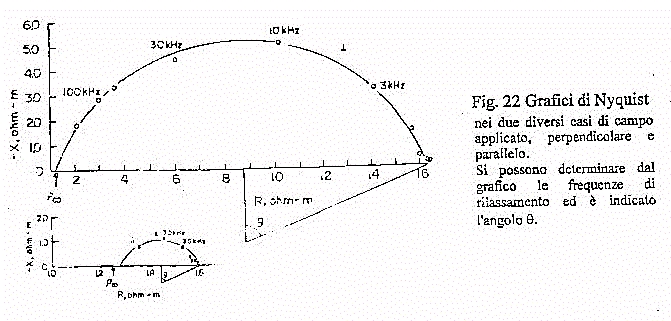 Fig.27
In figura 27 sono riportati i risultati sperimentali ottenuti da misurazioni
sul muscolo di un cane a 37 °C, sia nel caso di campo perpendicolare
che di campo parallelo. Si noti come nel secondo caso si abbia il rilassamento
intorno ai 12-13 kHz, un valore decisamente minore rispetto ai 70 kHz nel
caso di campo parallelo. Di contro si hanno valori di conducibilità
decisamente minori rispetto a quelli nella direzione ortogonale, il che
significa valori diversi per le perdite così come per la permettività.
Fig.27
In figura 27 sono riportati i risultati sperimentali ottenuti da misurazioni
sul muscolo di un cane a 37 °C, sia nel caso di campo perpendicolare
che di campo parallelo. Si noti come nel secondo caso si abbia il rilassamento
intorno ai 12-13 kHz, un valore decisamente minore rispetto ai 70 kHz nel
caso di campo parallelo. Di contro si hanno valori di conducibilità
decisamente minori rispetto a quelli nella direzione ortogonale, il che
significa valori diversi per le perdite così come per la permettività.
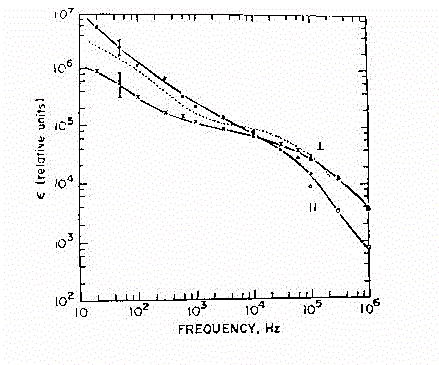 Fig.28
Fig.28
Si osservi ora la figura 28 nella quale e' illustrato l'andamento della
permittivita' nelle due condizioni di campo // e ^.
Si noti come a frequenze abbastanza basse ( ~50 Hz, frequenza della tensione
di rete), ci siano valori molto elevati di permettività e fortemente
dipendenti dalla direzione del campo applicato. A queste frequenze ciò
implica che un campo applicato su tessuto
muscolare genera una discontinuità all'interfaccia muscolo-aria.
Nel grafico la linea tratteggiata indica un andamento intermedio,
che è quello poi utilizzato come riferimento, visto che individuare
la direzione di campo all'interno del materiale non è sempre
facile. Ai valori medi ci si riferisce anche per "eliminare" la caratteristica
di anisotropia, considerando quindi il muscolo come un
tessuto misto.
2.4.4 - Dispersione beta
Nel caso del muscolo le non uniformità del materiale sono date
dal fatto che le cellule sono molto più lunghe in una direzione
che nell'altra. In questo range di frequenze le caratteristiche sono anche
più marcate Per fibre orientate parallelamente al campo, la dispersione
b è prodotta dalla carica della membrana
esterna, sebbene si presenti ad una frequenza minore di quelle audio22
proprio per la lunghezza della fibra. Nel caso di campo ^
la costante di tempo avrà un valore piuttosto contenuto.Generalmente
si suppone, effettuando una media sui valori, un raggio per le cellule
del muscolo di 50 mm ed una frazione di volume
delle cellule pari all'85%. Avremo i valori caratteristici:
es - e¥
@ aCm/e0
= 5*104
ss @
sa ( 1-r )/(
1+r ) = 0.06 [ S/m ]
( 2.5 )
fc @ si/2paCm
= 250 [ kHz ]
con:
si e sa
rilevati a 100 MHz; Cm = 10-2 F/m2.
Un'importante caratteristica dei tessuti si può poi dedurre
dalla figura 29: la variazione della temperatura (almeno per valori approssimativamente
all'interno del range del sistema termoregolatorio) non influenza in maniera
rilevante la permettività del materiale.
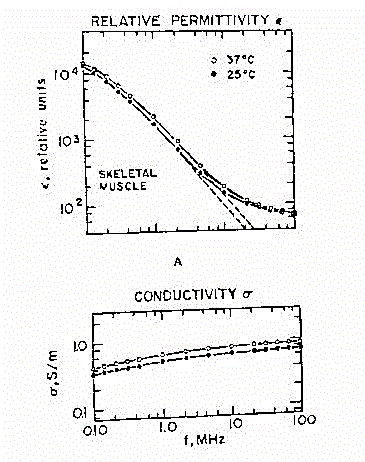 Fig.29
Fig.29
La figura 29 illustra la permittivita' e la conducibilita' del muscolo,
compensando l'anisotropia prendendo valori medi.Si fa notare soprattutto
il vistoso fenomeno di rilassamento che s'incontra intorno ai 250 kHz e
il valore asintotico, che si porta a circa 80, coincidente con il valore
della costante dielettrica dell'acqua. Ovviamente, salendo in frequenza,
i valori divergono leggermente, per la presenza delle proteine, dei sali
e delle cellule che si hanno nel materiale.
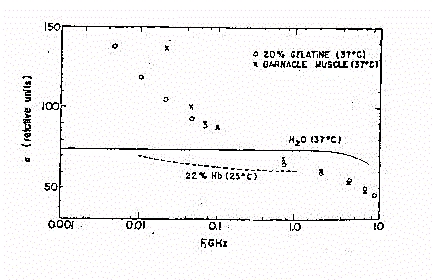 Fig.30
Fig.30
La figura 30 mostra comunque chiaramente il confronto tra l'andamento
della permettività nell'acqua ed in due diverse soluzioni (con la
medesima percentuale di cellule, in volume).
2.4.5 - Dispersione gamma
e delta
Per la dispersione di tipo g valgono, in
linea di massima, le considerazioni fatte nel caso dell'acqua.
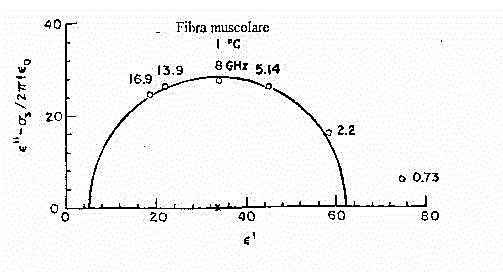 Fig.31
Nella figura 31 vediamo il diagramma di Nyquist relativo a misure su fibra
muscolare effettuate ad 1 °C. Come sappiamo, a temperatura minore corrisponde
una costante di tempo t maggiore, e di conseguenza
delle frequenze di rilassamento minori. Si noti come ad un intervallo di
variabilità per e' di ~57, corrisponde
un incremento di e'' pari a ~28. Il rilassamento
è localizzato intorno ad una fc
che va da 8 a 9 GHz, prossima quindi a quella dell'acqua pura (centrata,
si ricorda, sui 9 GHz).Ci sono però due incongruenze, una delle
quali osservabile anche dal grafico, che indicano la presenza di un ulteriore
rilassamento. I dati
Fig.31
Nella figura 31 vediamo il diagramma di Nyquist relativo a misure su fibra
muscolare effettuate ad 1 °C. Come sappiamo, a temperatura minore corrisponde
una costante di tempo t maggiore, e di conseguenza
delle frequenze di rilassamento minori. Si noti come ad un intervallo di
variabilità per e' di ~57, corrisponde
un incremento di e'' pari a ~28. Il rilassamento
è localizzato intorno ad una fc
che va da 8 a 9 GHz, prossima quindi a quella dell'acqua pura (centrata,
si ricorda, sui 9 GHz).Ci sono però due incongruenze, una delle
quali osservabile anche dal grafico, che indicano la presenza di un ulteriore
rilassamento. I dati
sperimentali, infatti, seguono abbastanza bene la curva dell'acqua
pura solo a partire da 2,2 GHz. Il valore a 0,73 GHz si
discosta notevolmente da quello supposto. Inoltre la frazione di volume
delle cellule a mobilità ridotta, calcolata tramite la teoria delle
misture è pari a 0,20, circa un quarto più di quello delle
proteine. Questo indica la presenza di una certa percentuale di
acqua con una frequenza di rilassamento minore. La restante parte d'acqua
(l'80 % in volume) segue invece l'andamento
dell'acqua pura. Questo strano comportamento, classificato come rilassamento
di tipo d è dovuto all'acqua legata.
C'è cioè una cospicua presenza di proteine che non ce la
fanno a seguire l'andamento del campo a causa della loro struttura. Queste
però sono molto cariche e tendono ad attirare le molecole d'acqua.
L'acqua si lega allora tramite legami elettrostatici ai punti di
carica sulla superficie delle proteine e s'incastra così in
questa specie di alveoli (detti "siti d'attivazione"). La proteina non
ha così tutti i suoi gradi di libertà e rilassa prima dei
2,2 GHz. Quanto la dispersione influisca sui valori della conducibilità
del tessuto
può essere visto nella figura 32, dove e' illustrata la conducibilita'
del tessuto muscolare per un campo perpendicolare, a due diverse temperature.
 Fig.32
L'informazione fornita non è dissimile da quella di figura 31. Anche
qui si fa vedere come, a 37 °C il tessuto presenti un ulteriore rilassamento
oltre quelli già noti b e g.
Il dato sperimentale al diminuire della frequenza indicato con un cerchietto
in figura è fortemente discorde dal dato teorico (estrapolato, come
indicato dalla linea tratteggiata). Si nota poi come queste differenze
diminuiscano notevolmente se si passa a temperature in cui l'acqua assume
una forma maggiormente strutturata, diminuendo cioè la temperatura
fino ad 1 °C. Le freccette indicano i valori di conducibilità
che si dovrebbe ottenere a 100 MHz alle due temperature di 1 °C e di
37 °C. Si noti come alle due diverse temperature, corrispondenti a
situazioni sperimentali differenti, si hanno due frequenze di rilassamento.
La cosa più rilevante è che il valore di conducibilità
riscontrato sperimentalmente ( ~6 S/m) in continua, è di gran lunga
maggiore rispetto a quello atteso ( ~1,5 S/m) a 100 MHz. Ciò significa
che la dispersione di tipo contribuisce in modo significativo alla conducibilità
totale. Possiamo poi vedere l'andamento delle proprietà dielettriche
al variare della temperatura (figura 33) in un diverso intervallo di valori.
Fig.32
L'informazione fornita non è dissimile da quella di figura 31. Anche
qui si fa vedere come, a 37 °C il tessuto presenti un ulteriore rilassamento
oltre quelli già noti b e g.
Il dato sperimentale al diminuire della frequenza indicato con un cerchietto
in figura è fortemente discorde dal dato teorico (estrapolato, come
indicato dalla linea tratteggiata). Si nota poi come queste differenze
diminuiscano notevolmente se si passa a temperature in cui l'acqua assume
una forma maggiormente strutturata, diminuendo cioè la temperatura
fino ad 1 °C. Le freccette indicano i valori di conducibilità
che si dovrebbe ottenere a 100 MHz alle due temperature di 1 °C e di
37 °C. Si noti come alle due diverse temperature, corrispondenti a
situazioni sperimentali differenti, si hanno due frequenze di rilassamento.
La cosa più rilevante è che il valore di conducibilità
riscontrato sperimentalmente ( ~6 S/m) in continua, è di gran lunga
maggiore rispetto a quello atteso ( ~1,5 S/m) a 100 MHz. Ciò significa
che la dispersione di tipo contribuisce in modo significativo alla conducibilità
totale. Possiamo poi vedere l'andamento delle proprietà dielettriche
al variare della temperatura (figura 33) in un diverso intervallo di valori.
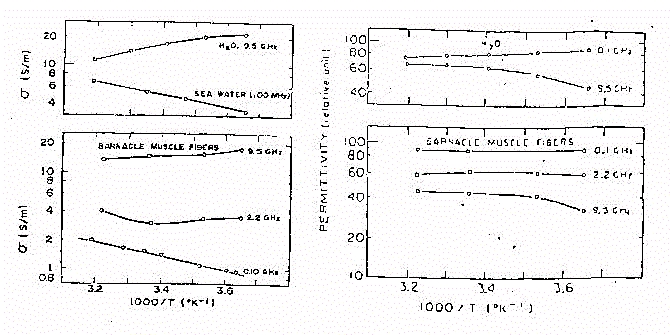 Fig 33
Nella figura 33 e' illustrato l'andamento delle caratteristiche dielettriche
del muscolo confrontate con soluzione saline e con acqua pura, al variare
di frequenza e temperatura. Fissata la frequenza, si hanno degli andamenti
più o meno costanti per s ed e
al variare della temperatura. Variazioni più intense si hanno invece
considerando delle frequenze differenti. Il che conferma l'impressione
già data dalla figura 32. La conducibilità intorno a 0,1
GHz ha lo stesso coefficiente di temperatura dei semplici "elettroliti".
A frequenze inferiori al rilassamento dell'acqua la conducibilità
che deriva dall'assorbimento dipolare varia con l'inverso di fc
e presenta un coefficiente di temperatura negativo pari a
circa -2% per °C. Quindi il coefficiente di temperatura della conducibilità
del tessuto decresce del 2% per grado alle frequenze dei GHz e ci sarà
un punto d'inversione solo a circa 2 GHz, quando contributi ionici e dipolare
sono uguali. I coefficienti di temperatura per la permettività sono
uguali o più piccoli. Il fatto che, comunque, a tutte le frequenze
non ci siano, né per e né per
s, variazioni più ampie del 2% per grado
(in senso positivo o negativo), le rende trascurabili nella maggiorparte
delle applicazioni pratiche.
Fig 33
Nella figura 33 e' illustrato l'andamento delle caratteristiche dielettriche
del muscolo confrontate con soluzione saline e con acqua pura, al variare
di frequenza e temperatura. Fissata la frequenza, si hanno degli andamenti
più o meno costanti per s ed e
al variare della temperatura. Variazioni più intense si hanno invece
considerando delle frequenze differenti. Il che conferma l'impressione
già data dalla figura 32. La conducibilità intorno a 0,1
GHz ha lo stesso coefficiente di temperatura dei semplici "elettroliti".
A frequenze inferiori al rilassamento dell'acqua la conducibilità
che deriva dall'assorbimento dipolare varia con l'inverso di fc
e presenta un coefficiente di temperatura negativo pari a
circa -2% per °C. Quindi il coefficiente di temperatura della conducibilità
del tessuto decresce del 2% per grado alle frequenze dei GHz e ci sarà
un punto d'inversione solo a circa 2 GHz, quando contributi ionici e dipolare
sono uguali. I coefficienti di temperatura per la permettività sono
uguali o più piccoli. Il fatto che, comunque, a tutte le frequenze
non ci siano, né per e né per
s, variazioni più ampie del 2% per grado
(in senso positivo o negativo), le rende trascurabili nella maggiorparte
delle applicazioni pratiche.
Note
21 - ATP, ovvero AdeninTriPhosfate
(adenintrifosfato), in cui all'adenina si legano tre atomi di fosforo.
Si scinde in due composti ADP ed AMP, rispettivamente adenindifosfato ed
adeninmonofosfato. Ad ogni scissione di un atomo di fosforo corrisponde
una liberazione d'energia molecolare; quasi tutti i processi biologici
si basano sulla capacità dell'organismo di effettuare il ciclo ATP-->
ADP--> AMP (processo catabolico, o meglio, metabolico)--> ADP-->ATP (processo
anabolico, ovvero l'assimilazione d'energia da processi come quelli digestivi,
ad esempio).
22 - Le frequenzeaudio sono quelle i cui
campi EM sono udibili dall'orecchio umano, ed ai sensi della nostra analisi
si considerano comprese tra20 e 200 kHz.