2.Materiali biologici
Indice Capitolo2
2.1 - Generalita'
La presenza dell'acqua nei tessuti biologici dà luogo a proprietàcaratteristiche
che possono essere messe in evidenza in vari
modi. Consideriamo ad esempio le conducibilità dei vari tessutibiologici
riportate in tabella; si può vedere come i valori siano fortemente
diversi tra materiale e materiale. Si vedrà poi come e quanto le
differenze dipendano proprio dalle diverse percentuali d'acqua contenute
nel materiale che si prende in considerazione (i valori sono considerati
ad una frequenza di 100 MHz e 3 GHz).
| f ( Mhz ) |
Muscolo |
Fegato |
Polmoni |
Milza |
Rene |
Materia bianca |
Materia grigia |
Osso |
Sangue |
Grasso |
| 100 |
0.9 -0.08 |
0.7 |
0.53 |
0.75 |
0.68 |
0.45 |
0.7 |
0.0574 |
1.0 |
0.02 |
| 3000 |
2.8 |
2.8 |
- |
2.7 |
2.3 |
1.5 |
2.0 |
0.16 |
2.7 |
0.04 |
Tab.7
L'importanza di quest'aspetto nello studio dell'interazione tra campi
elettromagnetici e tessuti biologici è messa poi in evidenza anche
dalle differenze che si riscontrano con misure tramite il modello a 180
celle utilizzato con la tecnica FDTD qualora le celle vengano considerate
omogenee per caratteristiche dielettriche, o vengano piuttosto valutate
con le loro effettive costanti, a seconda del materiale maggiormente presente
nella porzione di "corpo" simulato dalla cella.
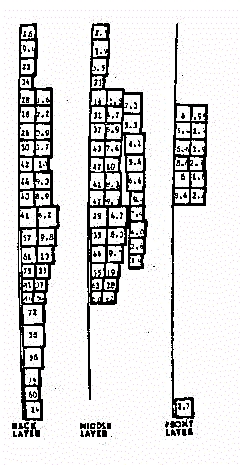
Fig.14 - Modello
a 180 celle disomogeneo
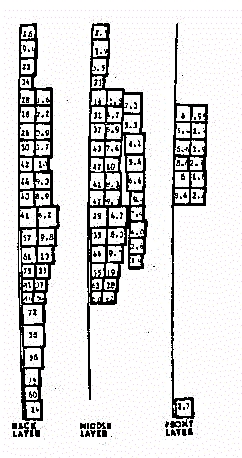
Fig. 15 -
Modello a 180 celle omogeneo
Nel primo modello si ha una migliore caratterizzazione dei tessuti
in quanto ad ogni cella vengono assegnate le caratteristichedielettriche
del materiale percentualmente più determinante. Si noti ad esempio
come ci siano maggiori dislivelli nei valori di SAR a livello della coscia,
dove, nella prima modellizzazione si e' tenuto conto della maggiore presenza
di tessutomuscolare, rispetto al resto della gamba, dove predomina la presenza
dell'osso.
Guardando i risultati sia sperimentali che tramite modelli si vede
come, aldilà di eventuali fenomeni di risonanza, la maggiore potenza
assorbita si ha nei tessuti a più alto contenuto d'acqua, secondo
lo schema di fondo :
CONTENUTO DI H2O Þ MAGGIORE CONDUCIBILITÀ
Þ MAGGIORE POTENZA DISSIPATA ALL'INTERNO.
2.2 - Proprietà dielettriche
dei materiali biologici
Procediamo dunque ad una suddivisione in tre gruppi principali. Tutti
i materiali all'interno di uno stesso gruppo sono caratterizzati da una
simile percentuale d'acqua e quindi l'andamento della permittività
e della conducibilità è analogo.
| Contenuto d'acqua |
Caratteristiche |
| Maggiore dell'80% |
In questa categoria sono inclusi tutti i fluidi con elettroliti, macromolecole
e materiale cellulare poco aggregato. Le cellule sono a grande distanza
e sono libere di muoversi nell'acqua che le circonda.
Esempi caratteristici sono: il fluido cerebrospinale, il sangue e l'umore
vitreo.
Abbiamo considerato l'occhio poiché il suo umore acqueo (contenuto
nella camera posteriore ed anteriore. Numero 3 in figura 16) è essenzialmente
una soluzione d'acqua e sali disciolti, e il corpo vitreo (numero 4 in
figura 16) è una massa "gelatinosa" composta da una particolare
soluzione salina. |
| Minore dell'80% |
In questo gruppo rientrano la pelle, i muscoli, la materia grigia e
la maggiorparte degli organi interni
(fegato, milza). Per i polmoni si ha un comportamento diverso poiché
contegono aria al loro interno. |
| Minore del 40% |
Sono fondamentalmente ossa, tendini e grasso caratterizzati da bassa
costante dielettrica, bassa
conducibilità e ridotto contenuto d'elettroliti.
Mentre per gli altri materiali non si apprezzano notevoli differenze
nelle caratteristiche e nella
composizione, al variare della specie animale da cui si prelevano i
campioni, per il tessuto grasso si hanno valori d'acqua che spaziano dal
2% fino a più del 40%.Otteniamo quindi valori per la costante dielettrica
e per la conducibilità diversi a seconda della specie considerata. |
Tab.8
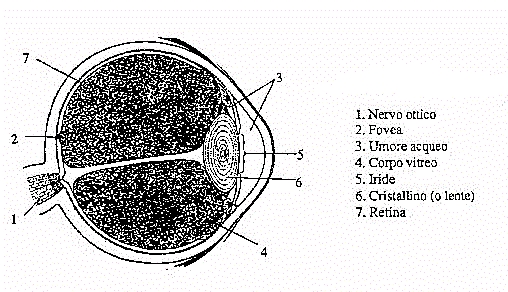 Fig.16 -
L' occhio umano
Fig.16 -
L' occhio umano
Prima d'analizzare nello specifico le caratteristiche di questi tre
gruppi, vediamo quali sono i diversi contributi in termini di e
e di s al variare della frequenza.
2.2.1 - A bassa frequenza
( fino a 100 khz )
La permettività puòraggiungere, in questo range di frequenze,
valori di 106. Si ha in questa zona la
presenza di tre fenomeni.
1- Il primo, ed il predominante, è l'effetto dovuto alla presenza
di elettroliti in soluzione (conducibilità ionica). Si ha spostamento
delle cariche,che muovendosi nel fluido extracellulare, e più raramente,
in quello intracellulare, danno un notevole contributo di perdite (vedi
figura 17) .
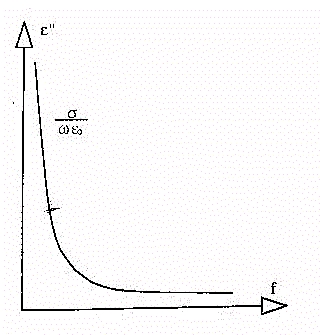 Fig.17 - Perdite da
conducibilita' ionica
Fig.17 - Perdite da
conducibilita' ionica
2- Si ha il contributo di rilassamento dovuto a polarizzazione da interfaccia.
Questo si verifica in soluzioni cellulari, ela frequenza di rilassamento
sarà funzione delle dimensioni delle cellule e dello spessore della
loro membrana. Usando la schematizzazione circuitale già analizzata16,
di un condensatore per la membrana, si ricorda che il fatto d'avere una
membranamolto sottile ed una grossa carica affacciata sulla superficie
dà un alto valore per la capacità, e di conseguenza, per
la e16.
3. Un terzo fenomeno è dovuto alla carica presente sulla superficie
della membrana che instaura una polarizzazione da controioni16.Questo
fenomeno è il primo che si realizza, ed è il maggiore responsabile
della diminuzione di e. Questo significa un
aumento di e",(una dispersione), e conseguentemente,
un rilassamento, che si definisce rilassamento di tipo a.
Ovviamente, a seconda del materiale, avremo diverse frequenze di rilassamento
(brevemente indicate con fc). La variazione
di conducibilità dovuta al rilassamento di tipo , deve confrontarsi
in questo range di frequenze, con l'andamento della conducibilità
trattato al primo punto.Supponendo una variazione complessiva De
di 106, ed una fc
~100 Hz, ci si aspetta un aumento di conducibilità (tramite le formuledi
Kramers-Kronig) pari a 0,005 S/m dovuto alla dispersione a.
I valori per la conducibilità ionica possono essere calcolati tramite
le formule di Maxwell-Fricke nel caso del sangue, il che è possibile
per l'ipotesi di una membrana molto sottile rispettoal volume occupato
dal fluido extracellulare. I calcoli sono stati effettuati utilizzando
un fattore di forma g variabile tra 0,65 ed1,39
(conseguenza delle diverse forme e dimensioni degli aggregati molecolari
nel sangue)17.
Considerando una conducibilità ionica per i normali fluidi biologici
pari a 2 S/m, ed una percentuale di fluido extracellulare per i tessuti
solidi dell'ordine del 10%, se ne ottiene un valore di conducibilità
alle basse frequenze di 0,14 S/m. Si vede come questi valori siano notevolmente
maggiori di quelli dovuti alla dispersione a,
rendendo questa dispersione di scarsa importanza dal punto di vista ingegneristico.
2.2.2 - A media frequenza
( fino a 300 Mhz )
Salendo in frequenza, la membrana, schematizzata con un condensatore,
si può considerare come cortocircuitata e si puo'
dunque applicare la teoria delle misture, considerando le proteine
contenute all'interno della cellula. Queste intervengono determinando il
cosiddetto rilassamento b. La differenza
rispetto alla dispersione di tipo a è
che l'effetto della conducibilità ionica è meno influente
di quella dovuta alla presenza di proteine nella soluzione. Questo perché
il valore di w è maggiore e quindi il
termine s/we diminuisce
d'importanza. In questo range di frequenze la curva della conducibilità
ionica presenta delle variazioni minime al variare della frequenza, e può
quindi quasi ritenersi costante. Per quanto riguarda e",
invece, sperimentalmente si nota che si ha una frequenza di rilassamento
fc intorno ai 500 kHz. Vediamo allora una
schematizzazione grafica dei vari contributi.
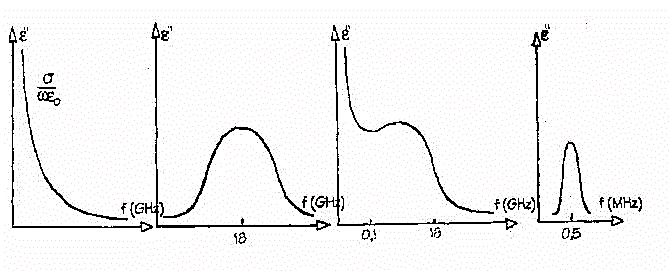 Fig.18
Nel primo grafico in figura 18 abbiamo riportato l'andamento della conducibilità
ionica, nel secondo l'andamento dell' acqua pura , nel terzo quello di
una soluzione salina, ottenuto come somma dei primi due. Il quarto grafico
rappresenta la dispersione di tipo b, che a
causa della bassa frequenza di taglio, è stata diagrammata su una
scala in MHz invece di una in GHz. Vediamo ora la composizione del terzo
e quarto grafico, che descrive l'andamento totale di una soluzione acquosa
alle medie frequenze.
Fig.18
Nel primo grafico in figura 18 abbiamo riportato l'andamento della conducibilità
ionica, nel secondo l'andamento dell' acqua pura , nel terzo quello di
una soluzione salina, ottenuto come somma dei primi due. Il quarto grafico
rappresenta la dispersione di tipo b, che a
causa della bassa frequenza di taglio, è stata diagrammata su una
scala in MHz invece di una in GHz. Vediamo ora la composizione del terzo
e quarto grafico, che descrive l'andamento totale di una soluzione acquosa
alle medie frequenze.
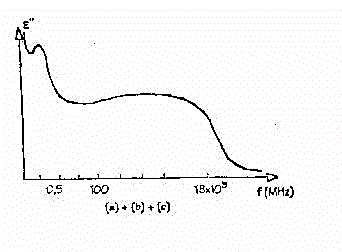 Fig.19 - Andamento complessivo
Fig.19 - Andamento complessivo
Si noti che questo grafico va ben oltre le medie frequenze. La prima
brusca variazione è dovuta al rilassamento di tipo b,
mentre il minimo relativo che si sperimenta intorno a 100 MHz è
dovuto all'effetto combinato della conducibilità ionica e di quella
dell'acqua pura. Il comportamento oltre queste frequenze è dovuto
al rilassamento dell'acqua pura.
Si nota che l'andamento intorno ai 100 MHz riporta una notevole flessione
del valore di e", al punto da presentare un
minimo. Questo perché fino a questa frequenza la conducibilità
ionica prevale sull'andamento dovuto all'acqua pura, facendo così
diminuire il valore complessivo di e". Oltre
il comportamento s'inverte, le perdite date dal rilassamento dell'acqua
sono maggiori e determinano l'andamento generale anche salendo in frequenza.
Inoltre si fa notare come, se non ci fosse il rilassamento di tipo
b, avremmo, alle medie frequenze, un andamento
pressoché costante, poiché, a queste frequenze, i due andamenti
si compensano.
2.2.3 - Ad alta frequenza
( UHF e m-onde , oltre i 100 Mhz )18
In questo range di frequenze abbiamo tre fenomeni:
1- Tra gli elettroliti in soluzione e le molecole di proteine, che
sono poco
conduttive, si genera una polarizzazione da interfaccia (alla Maxwell-Wagner).
E' particolarmente visibile solo in alcuni materiali, mentre generalmente
si hanno valori di poche centinaia di mS/m. La frequenza di rilassamento
che si sperimenta è comunque
intorno ai 300 MHz.
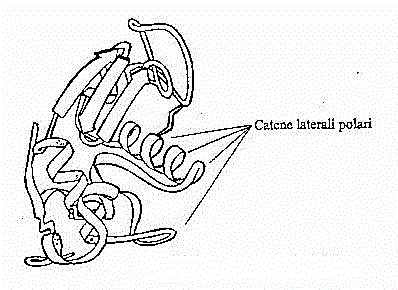 Fig.20 - Modello a
nastro di una proteina
2- Si possono avere perdite dovute a piccole molecole polari, come degli
amminoacidi in soluzione, oppure dovute alle catene laterali polari di
proteine (vedi figura 20). Avremmo, viste le minori dimensioni in gioco,
frequenze di rilassamento ben oltre i 100 MHz, dove si centra invece il
rilassamento dovuto alle intere catene proteiche. In realtà quest'ultimo
contributo determina l'andamento medio e la fc
rimane intorno ai 100 MHz.
Fig.20 - Modello a
nastro di una proteina
2- Si possono avere perdite dovute a piccole molecole polari, come degli
amminoacidi in soluzione, oppure dovute alle catene laterali polari di
proteine (vedi figura 20). Avremmo, viste le minori dimensioni in gioco,
frequenze di rilassamento ben oltre i 100 MHz, dove si centra invece il
rilassamento dovuto alle intere catene proteiche. In realtà quest'ultimo
contributo determina l'andamento medio e la fc
rimane intorno ai 100 MHz.
3- Abbiamo poi il contributo dovuto al rilassamento dell'acqua pura,
caratterizzato da un processo che presenta una singola
costante di tempo t pari ad 8 psec, a cui
corrisponde una frequenza di rilassamento pari a ~20 GHz a 25 °C.
Per frequenze f << fc possiamo
assumere, per la conducibilità s, una
dipendenza dal quadrato della frequenza. Si veda la formula:
s = [2pf2
( es-e¥
)e0 / fc ] / [1+ ( f/fc
)2]
(2.2)
Alle basse frequenze il denominatore si approssima ad uno, e rimane la
sola dipendenza quadratica del numeratore. L'andamento asintotico tende
invece ad un valore costante, come prevedibile da considerazioni fisiche
sulla mobilità. Per tessuti ad elevato contenuto d'acqua, nel range
di frequenze tra 3 e 5 GHz, quest'aumento di conducibilità è
dello stesso ordine di grandezza di quella ionica. Questa compensazione
genera un andamento costante nell'intervallo di frequenza (vedi figura
19). Inoltre la conducibilità dei tessuti e delle soluzioni proteiche,
includerà il contributo del rilassamento dipolare dell'acqua legata,
indicato come rilassamento di tipo d,
con una fc di circa un ordine di grandezza
minore di quella dell'acqua libera. La conducibilità totale è
dunque definita da tre contributi:
- conducibilità ionica;
- rilassamento dovuto all'acqua legata;
- rilassamento dovuto all'acqua libera.
Sperimentalmente, però, sono tutti compresi tra 3 e 5 GHz, e
non è sempre semplice separare le singole componenti.
Complessivamente possiamo osservare le caratteristiche dei tessuti
con acqua comparando i dati sperimentali con quelli previsti nel caso di
un rilassamento modellizzato con una sola costante di tempo. Si può
rappresentare la conducibilità come la somma
di un termine dipolare e di uno ionico:
s = [2pf2
( es-e¥
)e0 / fc ] / [1+ ( f/fc
)2] + ss
(2.3)
in cui i parametri es,e¥
ed fc si riferiscono all'acqua pura (contributo
dipolare) e ss
alla conducibilità ionica. Supponendo le proteine non conduttive,
possiamo riscrivere l'equazione per la conducibilità ricavata dalla
teoria delle misture, nel caso di si
<< sa:
s ® ( 1-r'
)sa / ( 1+r'/2
) + 9r'si
/ ( 2+r' )2
(2.4)
dove si è
il contributo di conducibilità del sistema proteine-acqua legata,
e p' è la frazione di volume delle proteine disciolte in soluzione.
Questa frazione è maggiore di p (frazione di volume delle proteine
non idratate). La differenza (p' - p) indica la percentuale d'acqua a mobilità
ridotta. Se si sceglie una frequenza sufficientemente elevata, le variazioni
di si
diventano
trascurabili, e la dipendenza di s dalla
frequenza riflette quella di sa.
In sa il contributo
ionico è a sua volta trascurabile rispetto a quello dipolare, e
ci confrontiamo ancora una volta con la prima equazione scritta per s,
ovvero una funzione lineare di f2 / [1
+ (f / fc)2]
qualora fc sia quella della maggiorparte
del liquido. Dopo questa discussione sui fenomeni che intervengono alle
varie frequenze, analizziamo ora le caratteristiche dielettriche dei tre
gruppi di tessuti che si erano prima distinti.
Note
16 - Stiamo cioè considerando una
situazione del tipo:
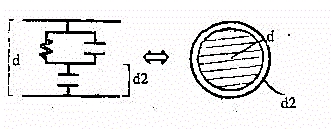 Fig.21
Fig.21
Considerando il materiale interno dissipativo e quelo esterno non conduttivo,
alle basse frequenze si avrà per la permittività la formula:
es
= e2d / d2
( 2.1 )
17 - La teoria utilizzata mantiene la sua
validità anche per tessuti solidi, ma i risultati sono di più
difficile predizione a causa della
maggiore complessità strutturale. Viene comunque simulata sufficientemente
bene con la teoria delle misture.
18 - Si noti come le fasce in cui abbiamo
suddiviso le frequenze non siano troppo stringenti, e si possa quindi considerare
alte frequenze già i 100 MHz.