CENNI SUL SISTEMA NERVOSO UMANO
Le unità che compongono il sistema nervoso sono
le cellule nervose o neuroni. 
La struttura base di ogni neurone comprende una membrana,
il corpo cellulare, o soma, e speciali prolungamenti che si dipartono dal
contorno del soma portandosi a varia distanza da esso. Queste propaggini
sono distinte in dendriti in genere molti e ramificati e un unico assone
che, lungo il suo decorso, costituisce la fibra nervosa.
L' assone è rivestito da una membrana e contiene,
all'interno, l'assoplasma gli assoni di diversi neuroni concorrono a formare
fasci di fibre o nervi.
Nel corpo cellulare del neurone, avviene la sintesi di
alcune sostanze che svolgono una funzione di trasmissione chimica degli
impulsi nervosi tra neuroni e tra neurone e fibra muscolare.
Nelle fibre nervose amieliniche, gli assoni sono racchiusi
semplicemente in una cellula di Schwann. Invece nelle fibre mieliniche
l' assone è ulteriormente circondato da una guaina mielinica che
è una serie specializzata di cellule di Schwann avvolte ripetutamente
intorno all'assone.
La guaina mielinica manca alla terminazione dell' assone
e a livello di periodiche costrizioni, distanti tra loro circa 1 mm, dette
nodi di Ranvier. L'assone termina in un numero di bottoni sinaptici, o
sinapsi, che contengono delle vescicole nelle quali si deposita il mediatore
chimico secreto dal nervo.
Le sinapsi sono collegate al corpo cellulare o ai dendriti
di un altro neurone. Un neurone può trovarsi in uno stato quiescente
o eccitato. Nel secondo caso, lungo l' assone vengono trasmessi dei segnali
ad altri neuroni tramite le sinapsi, che a loro volta possono essere eccitatorie
o inibitorie nei confronti dei neuroni a cui trasmettono il segnale. Gli
assoni che terminano nei muscoli si congiungono alle fibre muscolari mediante
una placca motrice.
Molto schematicamente si può dire che il sistema
nervoso è una struttura nella quale reti e catene neuroniche, che
assicurano connessioni molteplici ed elevatissime possibilità di
convergenza e divergenza degli impulsi su singole unità o su gruppi
di neuroni, sono in grado di ricevere e talora immagazzinare nell' unità
di tempo un numero ragguardevole di informazioni e sono capaci di elaborare
per alcune informazioni ricevute risposte più o meno complesse.
Il sistema nervoso si può dividere in tre parti: centrale, periferico
e autonomo. Il sistema nervoso centrale è formato da una struttura
tubolare che varia nello spessore e nella forma in differenti regioni.
Esso può essere a sua volta suddiviso in tre porzioni: midollo spinale,
tronco cerebrale ed encefalo.
Al midollo spinale pervengono informazioni dall' interno
o dall'esterno dell' organismo; esso elabora risposte relativamente elementari
in rapporto agli stimoli ricevuti.
Nel tronco cerebrale vengono coordinate informazioni
più complesse come quelle provenienti dalle vie uditive e visive.
Nell' encefalo si raggiunge il grado più elevato
di elaborazione delle informazioni afferenti, così come è
possibile elaborare risposte motrici di grande complessità, nonché
svolgere tutte quelle attività che caratterizzano la vita di relazione.
Il sistema nervoso periferico è formato dai gangli
e dai nervi che connettono la parte centrale del sistema nervoso con gli
organi di senso e gli organi effettori, quali i muscoli e le ghiandole.
I nervi di senso, più propriamente detti fibre
afferenti, inviano al sistema nervoso centrale gli impulsi raccolti dalla
periferia. In particolare la cute è riccamente innervata di recettori,
compresi quelli del dolore. I nervi motori, detti fibre efferenti, trasportano
gli impulsi che vanno dal sistema nrrvoso centrale ai muscoli volontari.
Il sistema nervoso autonomo o simpatico opera in modo
involontario e controlla molte funzioni. Si divide nei sistemi ortosimpatico
e parasimpatico, che sono ad azione antagonista nel senso che controllano
effetti opposti nei vari organi: il livello di normale attività
è determinato dall' equilibrio dei due sistemi. Analizziamo in dettaglio
il sistema nervoso centrale.
Il midollo spinale, le cui pareti spesse sono
costituite da migliaia di fibre nervose, è protetto dalle prime
ventidue vertebre della colonna vertebrale. Attraverso una sezione trasversale
si possono vedere due diversi tipi di materiale. Al centro è presente
una massa a forma di H di sostanza -grigia-, costituita dai corpi cellulari
dei neuroni. Intorno a questa si ha della sostanza bianca formata da fibre
mieliniche riunite in fasci. La massa ad H può essere vista come
formata da coppie di corna posteriori ed anteriori. Dalle corna
posteriori escono le fibre afferenti, mentre da quelle anteriori escono
le fibre efferenti. Vi sono poi dei fasci di fibre che collegano il midollo
spinale con vari centri del cervello. In tal modo gli impulsi provenienti
dai recettori arrivano al cervello, il quale può, a sua volta, inviare
impulsi ai neuroni motori e quindi ai muscoli.
Tutti gli impulsi afferenti provenienti dall periferia
penetrano nel midollo spinale attraverso le cosiddette radici dorsali.
Nel sistema nervoso centrale e nelle radici dorsali sono numerose le fibre
amieliniche.
Il cervello può anche non intervenire, come nelle
azioni riflesse. Ciò avviene attraverso un circuito nervoso detto
arco riflesso, consistente in un collegamento tra fibre afferenti
ed efferenti all'interno della materia bianca del midollo spinale.
Il tronco cerebrale possiede circuiti riflessi
simili a quelli del midollo spinale, ma sovrapposti a questi troviamo circuiti
integrativi di attività che interessano l'organismo come un tutto.
Il tronco cerebrale svolge qindi la funzione di centro di collegamento
e di smistamento degli impulsi che arrivano e partono dai centri superiori.
Il cervello, protetto dal cranio e dallo scalpo,
è la parte principale e più cospicua dell'encefalo. La superficie,
che distesa avrebbe un'area di circa 1 m^2, è percorsa da solchi
e circonvoluzioni caratteristiche per ogni individuo. All'interno del cranio
sono presenti diverse sostanze in proporzioni diverse, le più abbondanti
delle quali sono la materia bianca, la materia grigia ed
il fluido cerebre spinale.
Il cervello è parzialmente diviso negli emisferi
destro e sinistro. In contrasto con la quasi perfetta simmetria
bilaterale che il cervello mostra all'esame anatomico, esiste un'ineguale
distribuzione delle funzioni tra i due emisferi. Inoltre, l'area corticale
motoria dell'emisfero sinistro corrisponde alla parte destra del corpo,
e viceversa. I due emisferi sono collegati funzionalmente per mezzo delle
fibre commissurali.
La volta del cervello è composta di sostanza grigia
ed è detta corteccia cerebrale. Essa è caratterizzata
da un gran numero di connessioni tra le fibre afferenti provenienti dalle
aree sensoriali del cervello e dal midollo spinale e tra queste e le fibre
efferenti La corteccia è la principale area di associazione ed è
composta da una stratificazione di cellule nervose essenzialmente di due
tipi: le cellule piramidali e quelle stellate, disposte in
sei strati. Oltre alle fibre afferenti ed efferenti e alle fibre di associazione
che connettono aree diverse di uno stesso emisfero, esistono anche fasci
di fibre che connettono i due emisferi, come il corpo calloso. La maggior
parte dei neuroni della corteccia e i grandi nervi periferici sono mielinizzati.
In ciascuno degli emisferi è possibile distinguere
quattro lobi. I lobi frontali presiedono alle attività
di pensiero, della parola ed al controllo dei movimenti. I parietali
sono relativi alla sensibilità cutanea alla pressione e alla
temperatura, gli occipitali alla vista e i temporali al gusto, all'odorato
e all'udito. Stimolando opportunamente alcune zone della corteccia si possono
attivare le funzioni associate.
Il cervelletto è situato sotto la sezione
posteriore del cervello ed è interconnesso in maniera complessa
con le altre parti del sistema nervoso. Esso regola e coordina l'attività
motrice dell'intero organismo, col mantenimento dell'equilibrio del corpo
e intervenendo nell'esecuzione di movimenti che richiedono una grande precisione.
Modello di fibra nervosa
All' interno del singolo neurone l' informazione
viaggia sotto forma di impulso elettrico: per comprendere come ciò
avvenga bisogna considerare le proprietà della membrana cellulare.
La membrana cellulare è costituita da un doppio
strato di fosfolipidi intervallato da proteine. La matrice fosfolipidica
è praticamente impermeabile agli ioni i quali possono attraversare
la membrana solo in corrispondenza alle proteine le quali funzionano quindi
come dei veri e propri canali.
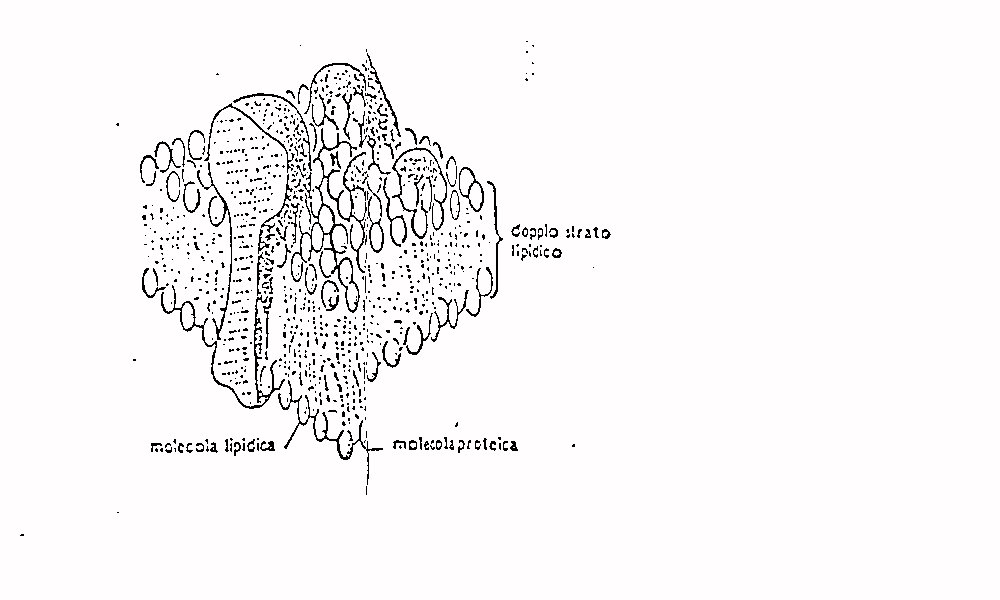
Fig . 4
Le proteine sono delle macromolecole biologiche formate
da sequenze di amminoacidi e possono assumere differenti configurazioni
nello spazio sotto l'influenza di agenti esterni quali : calore, campo
elettrico, energia luminosa o presenza di altri composti .
Dato che le proteine presenti nella membrana di una cellula
funzionano come dei canali per gli ioni, una variazione della forma di
queste proteine determinerà una variazione nella permeabilità
dei canali.
In particolare consideriamo i canali a controllo di ligando
( come ad esempio i canali del calcio nelle fibre muscolari in prossimità
della placca neuromotoria ) che si aprono quando un ligando extracellulare
( ad esempio l'acetilcolina ) si lega alle proteine del canale ed i canali
a controllo di potenziale che si aprono quando la differenza di potenziale
ai capi della membrana raggiunge un valore di soglia.
Le proteine della membrana cellulare, oltre che come
canali, possono funzionare anche come pompe permettendo di effettuare un
trasporto di materia attraverso la membrana in verso contrario al gradiente
elettrochimico a spese del metabolismo della cellula; di fondamentale importanza
è la pompa sodio potassio che permette di avere all'interno della
cellula una bassa concentrazione di sodio ed una elevata concentrazione
di potassio.
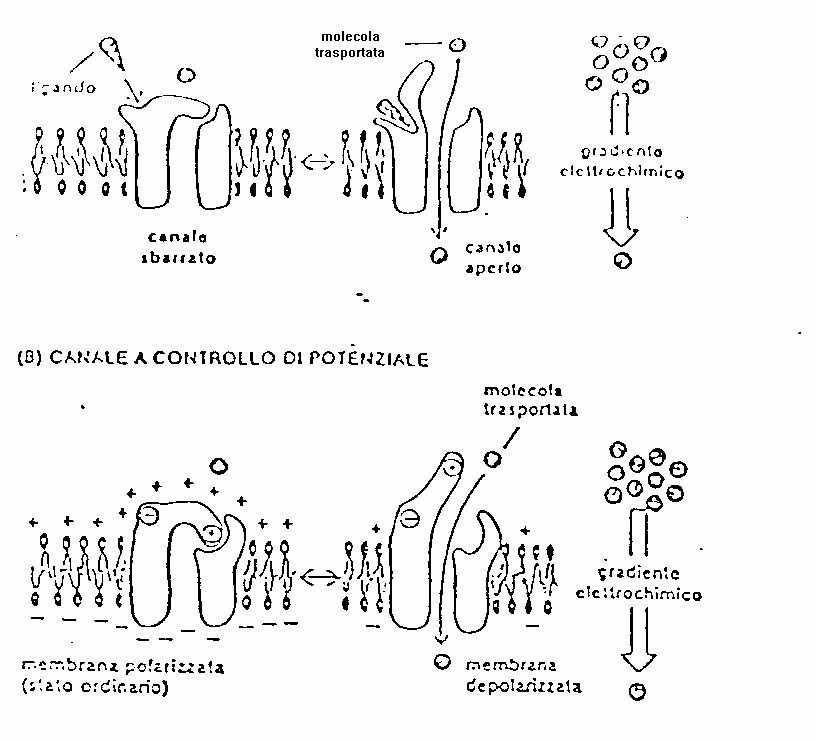
Fig . 5
La presenza di concentrazioni di ioni diverse ai
due lati della membrana determina una differenza di potenziale fra l'interno
e l'esterno della cellula.
Per un neurone a riposo questa differenza è di
circa 85 mVolt con l'interno più negativo dell'esterno.
L'impulso nervoso non è altro che una variazione
di questa differenza di potenziale che si propaga lungo la fibra ed ha
un andamento nel tempo come quello riportato in figura 6.
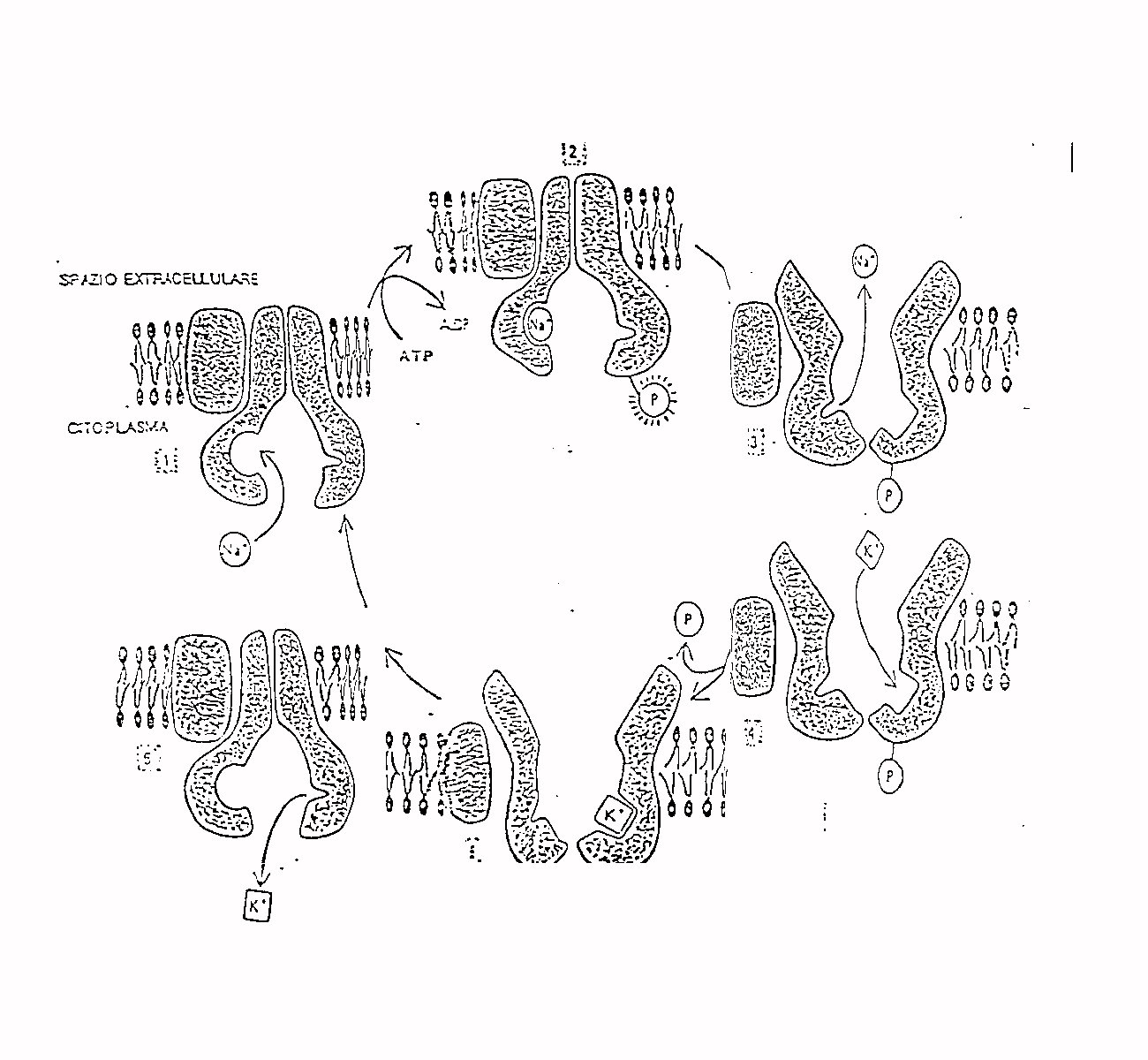
Fig . 6
L'impulso nervoso viene con più precisione definito
potenziale d'azione: una descrizione quantitativa di questo fenomeno
è fornita dal modello di Hodgkin & Huxley.
Consideriamo innanzitutto l'equazione di Goldman per
una singola specie ionica :
( 12 ) 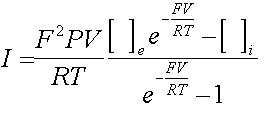
Dove I è la corrente che scorre attraverso
la membrana, F è la costante di Faraday, V è la differenza
di potenziale ai capi della membrana, P è la permeabilità
della membrana per la specie ionica considerata, R è la costante
dei gas perfetti, T è la temperatura in gradi kelvin, [ ]e è
la concentrazione esterna della specie ionica e [ ]i la concentrazione
interna. Tenendo conto che la membrana ha anche un comportamento capacitivo
possiamo modellarla con un circuito equivalente come quello di figura 7,
dove abbiamo considerato i contributi del sodio del potassio e del cloro
ma in cui non compare esplicitamente l'effetto della pompa sodio potassio
che può essere modellata con un generatore di corrente. C'è
da notare che i valori delle conduttanze dipendono in maniera non lineare
da V.
Un circuito più sofisticato che tiene conto anche
del contributo del calcio e quello di figura 8
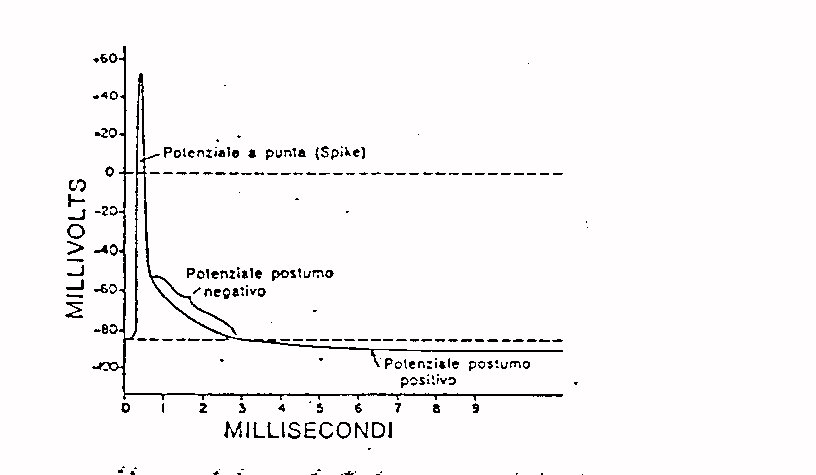
Fig . 7
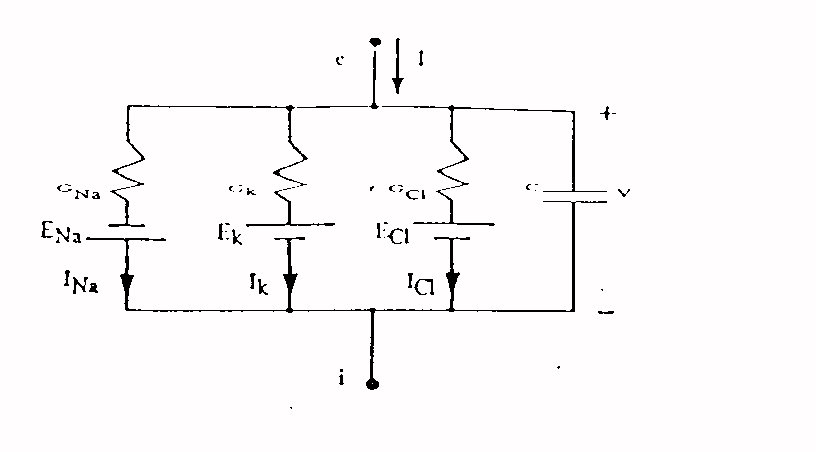
Fig . 8
Hodgkin & Huxley misurarono l'andamento delle conducibilità
per il sodio e per il potassio in funzione del voltaggio ai capi della
membrana. Questi andamenti furono ricavati usando la tecnica del voltage
clamp e quindi in condizioni sperimentali molto diverse da quelle fisiologiche
ma approssimano in maniera soddisfacente la situazione reale.
I risultati possono essere espressi dalle seguenti relazioni:
_
( 13 ) 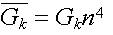
_
dove Gk è una costante con dimensioni conduttanza
/ superficie e n è dato da
( 14 ) 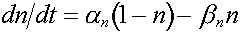
dove an e bn dipendono solo dal
voltaggio V ai capi della membrana ed hanno la seguente espressione
( 15 ) 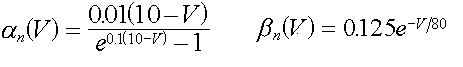
Per la conduttanza relativa al sodio valgono delle
formule analoghe
(16-17-18-19)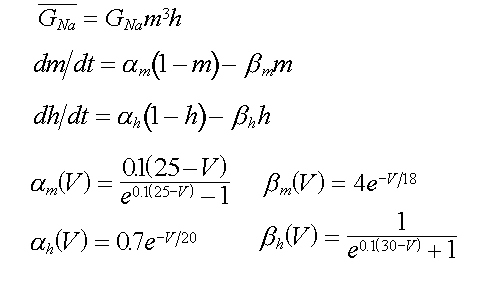

Fig . 9
Le figure 9 e 10 mostrano landamento nel tempo delle
conduttanze relative al potassio ed al sodio in risposta ad un potenziale
a gradino ( le diverse curve sono per gradini di diversa ampiezza )
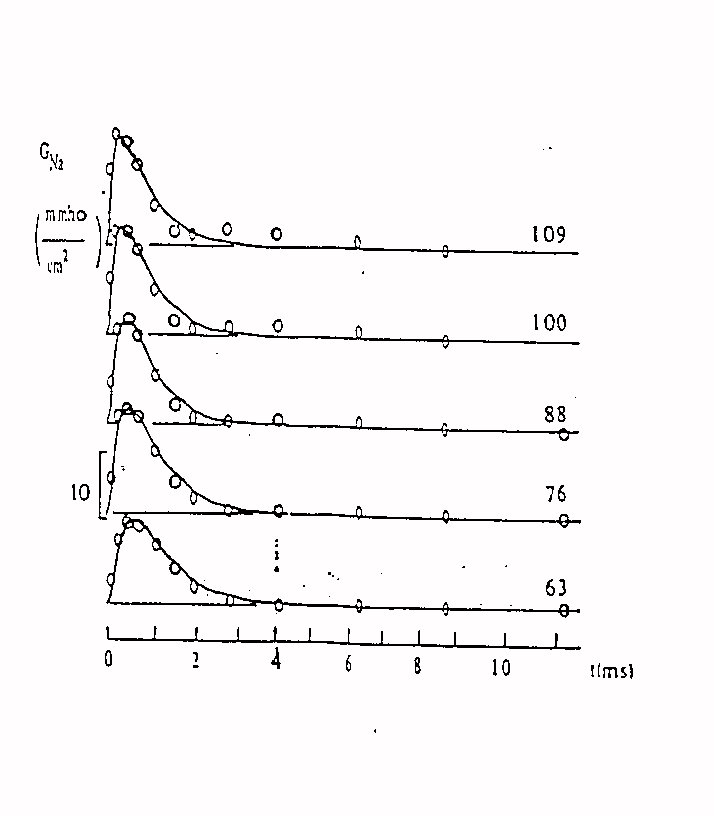
Fig . 10
Possiamo cercare di interpretare il modello dicendo che
se la depolarizzazione raggiunge un livello sufficiente, detto livello
di soglia, si instaura un processo autorigenerativo che possiamo descrivere
così:
1) il potenziale di membrana diminuisce,
2) la permeabilità al sodio aumenta di circa 500
volte,
3) il sodio presente nel liquido extracellulare si riversa
all'interno della cellula,
4) con l'entrata di sodio aumenta la carica positiva all'interno
della cellula e quindi il potenziale di membrana diminuisce ancora di più.
Questo processo continua fino a che non si raggiunge
una situazione di inversione della polarizzazione in cui l'interno della
cellula è più positivo di 45 mV rispetto all'esterno, il
che avviene in circa 0.5 ms. A questo punto la permeabilità per
il sodio ritorna ai valori normali e aumenta invece quella per il potassio
che quindi fuoriesce dalla cellula fino a che il potenziale non sia ritornato
al valore di riposo, contemporaneamente anche la permeabilità per
il potassio diminuisce gradualmente fino al suo valore di riposo. Sarà
infine compito delle pompe ristabilire la concentrazione dei due ioni.
Possiamo inoltre notare che uno stimolo tale da provocare
una depolarizzazione che non raggiunga il livello di soglia non darà
luogo ad alcun impulso mentre uno stimolo bastevole ad originare una depolarizzazione
che raggiunga il livello di soglia darà origine ad un impulso di
ampiezza costante, determinata solo dalle caratteristiche della membrana
ed indipendente dall'intensità dello stimolo: questo è ciò
che va sotto il nome di legge del tutto o nulla.
Perché ci sia una trasmissione dell'impulso nervoso
il potenziale d'azione deve propagarsi lungo la membrana: ciò avviene
in maniera diversa nelle fibre amieliniche ed in quelle mieliniche.
Nelle fibre amieliniche il passaggio di ioni può
avvenire in qualunque punto della membrana; quando si genera un potenziale
di azione in una porzione di membrana questo depolarizza le zone di membrana
adiacenti e da' origine a dei nuovi potenziali d'azione che si propagano
in tutte le direzioni.
Nelle fibre mieliniche invece l'assone è circondato
da una guaina mielinica formata da cellule dette cellule di Schwann che
si avvolgono a spirale e formano uno strato isolante; questo strato si
assottiglia in corrispondenza della giunzione fra soma e assone, in corrispondenza
delle terminazioni sinaptiche e si assottiglia anche in corrispondenza
a di punti posti periodicamente lungo l'assone a distanza di circa 2 mm
l' uno dall'altro detti nodi di Ranvier.
Sono questi gli unici punti della membrana attraverso
cui gli ioni possono fluire e dove quindi si può instaurare un potenziale
d'azione.
Nelle fibre mieliniche quindi il potenziale d'azione
salta da un nodo di Ranvier all'altro ed a questo fenomeno si da appunto
il nome di conduzione saltatoria.
Da quanto detto appare chiaro come l'impulso viaggerà
più velocemente nelle fibre mieliniche, in particolare la velocità
di conduzione può variare nelluomo da 0.5 m / s per le più
piccole fibre amieliniche fino a 130 m / s per le più grandi fibre
mieliniche.
Le fibre amieliniche si trovano principalmente nel sistema
nervoso centrale mentre quelle mieliniche formano la maggior parte dei
nervi periferici ed è quindi a queste ultime che siamo interessati.
Useremo il modello di Mc Neal in cui si usa un circuito
equivalente come quello di Hodgkin & Huxley per rappresentare un nodo
di Ranvier ed i vari nodi sono collegati da una conduttanza che rappresenta
il liquido intracellulare:
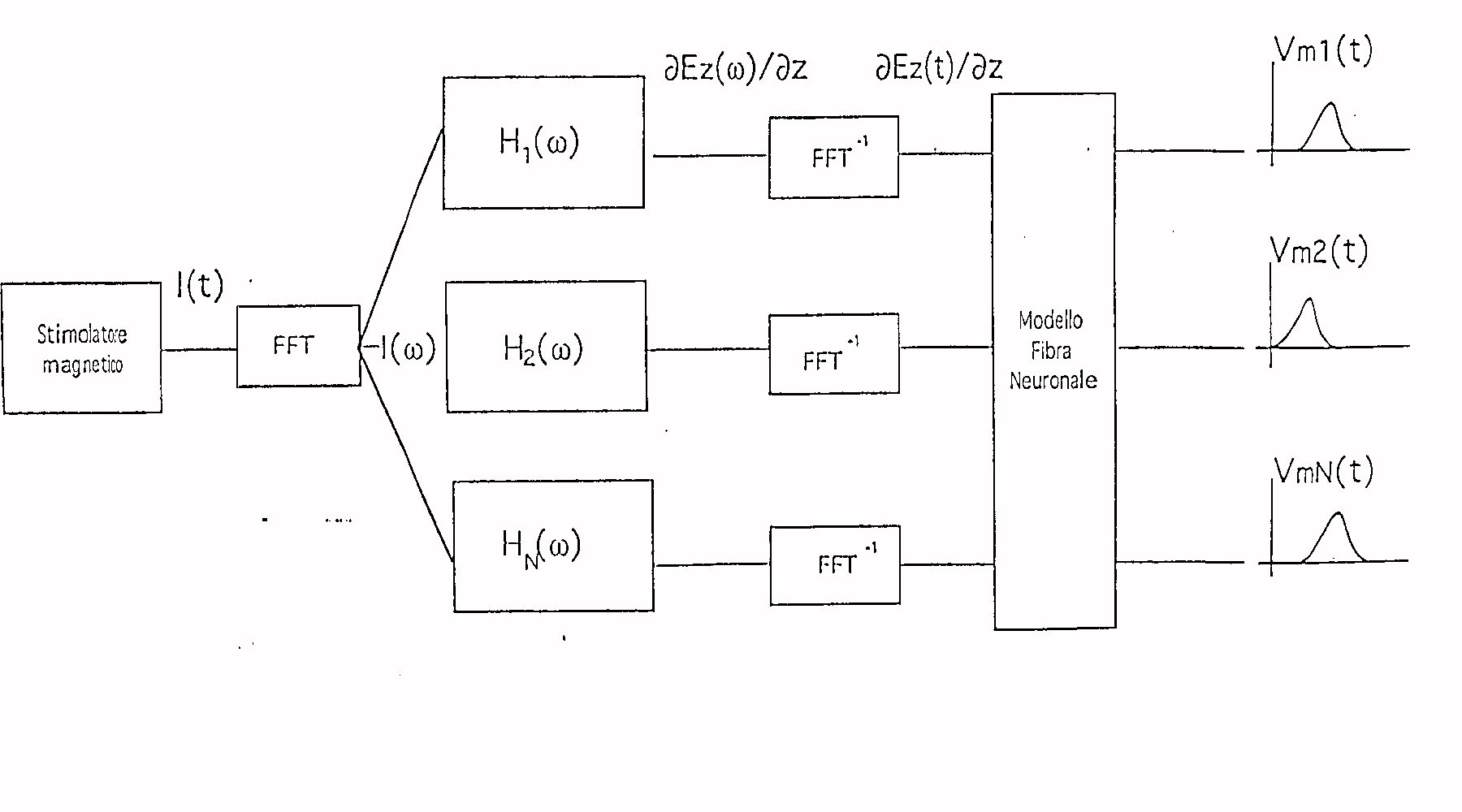
Fig . 11
METODI
Lo scopo del lavoro consiste nello sviluppare un
metodo che consenta , a partire dalle caratteristiche dello stimolatore,
di valutare la dinamica della eccitazione nervosa. L' approccio seguito
fa riferimento allo schema di principio di fig.
Nel blocco relativo allo stimolatore rientra la bobina
con le sue caratteristiche, la quale è percorsa dalla corrente I
( t ) . Di questa corrente si fa la FFT ottenendo la I ( w ) che
viene inviata al blocco H1( w ) nel quale applicando il MEAM si calcola
la derivata spaziale lungo la direzione z della componente del campo elettrico
lungo la medesima direzione all' interno del soggetto.
A questo punto si antitrasforma ed il risultato viene
inviato in ingresso al blocco relativo al modello della fibra neuronale
la cui uscita ci permette di rilevare la eventuale generazione di un potenziale
d' azione.
L' utilità di questo schema risiede nel fatto
che esso ci permette di valutare l' influenze che i diversi fattori ( come
l' intensità della corrente I( t ) e la forma della bobina
) hanno sulla generazione del potenziale d' azione.
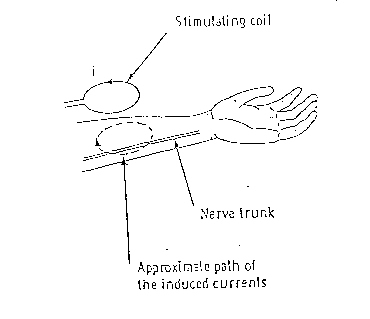
Fig . 12